Кто выдвинул идею химической эволюции. Основные этапы химической и биологической эволюции
Химическая эволюция.
Химическая эволюция:
начальные этапы.
Центральные части
Солнца и других звезд почти
не имеют в своем составе настоящих
химических элементов и образованы в основном
из плазмы. Плазма - полностью ионизированный
газ, состоящий из хаотически движущихся
положительно заряженных (атомные ядра)
и отрицательно заряженных (электроны)
частиц.
Строение вещества звезд
определяется степенью ионизации (процентом
вещества, находящегося в состоянии плазмы).
В центральной части Солнца температура
составляет от 3 до 20 млн. градусов. При
этой температуре степень ионизации достигает
100%, т.е. все вещества находятся в состоянии
плазмы. На глубине, равной 0,1 радиуса Солнца,
температура снижается до 400 000* С, а на
поверхности Солнца температура падает
до 5500* С. При этом степень ионизации снижается
до 0,01%, т.е. 99,99% веществ на поверхности
Солнца находится в виде атомов, имеющих
электронные оболочки.
Спектральными анализами
на поверхности Солнца обнаружено около
60 химических элементов, среди которых
преобладают водород и гелий. Это объясняется
тем, что другие элементы с более высокой
атомной массой и более сложной структурой
атомного ядра и электронной оболочки
не могут долго существовать при высокой
температуре. Количество атомов водорода
в солнечной атмосфере в 4-5 раз больше
количества атомов гелия; количество атомов
всех других элементов в 1000 раз меньше
количества водорода.
В глубинах Солнца и звезд,
в плазме происходит образование
сложных ядер из простейших вследствие
захвата протонов и нейтронов. Образование
ядра гелия из водорода идет в три
этапа. Из ядра водорода (протона) и
нейтрона образуется ядро тяжелого водорода
(дейтерия - D) - дейтрон. При соединении
дейтрона с еще одним протоном
образуется ядро легкого изотопа
гелия - Не|. В результате слияния двух
ядер легкого гелия образуется ядро обычного,
тяжелого гелия - Не2 и высвобождается
два протона.
В ходе термоядерных реакций
создаются ядра новых элементов.
При соединении трех ядер гелия возникает
ядро изотопа углерода.
В результате присоединения
к ядру углерода других частиц гелия
возникают изотопы кислорода, неона,
магния и других элементов. Таким
образом, возникновение атомов химических
элементов - начальный этап неорганической
эволюции. Водород, углерод, кислород,
азот, фосфор (так называемые биогенные
элементы) широко распространены в космосе
и имели большую возможность реагировать
между собой с образованием простейших
неорганических соединений - следующий
этап неорганической эволюции. Этому способствовало
наличие энергии в космосе в виде электромагнитного
излучения и тепла, испускаемого звездами.
Преобладание водорода, кислорода, азота
и фосфора в живых системах не случайно:
водород - хороший восстановитель, легко
образует с кислородом и азотом водородные
связи, имеющие большое значение в образовании
биологических структур и для процессов
жизнедеятельности. Кислород обладает
большой окислительной активностью, а
для фосфора характерно образование макроэргических
связей, в которых запасается энергия
при химических реакциях.
Третий этап химической эволюции
- образование простейших органических
соединений - связан со специфической
валентностью углерода - главного носителя
органической жизни, его способностью
к соединению почти со всеми элементами,
к образованию цепей и циклов, с его каталитической
активностью и другими свойствами. Простейшие
органические молекулы широко распространены
в межзвездной среде.
Первый этап химической
эволюции на Земле.
Химическая эволюция -
это совокупность процессов, протекавших
в Космосе и на ранних этапах существования
Земли, приведших к возникновению жизни.
На первом этапе образовались литосфера,
гидросфера, атмосфера. Литосфера возникла
вследствие вулканизма. Ежегодно вулканы
выбрасывают на поверхность Земли около
1 км. За время существования Земли, при
нынешней активности вулканов, было выброшено
такое количество лавы, которой достаточно
для образования коры Земли.
Гидросфера также создана
вулканами: 3 % массы лавы составляет
водяной пар. Пар конденсировался.
Это привело к появлению осадков
и Первичного океана. Атмосфера образовалась
при дегазации лав. Вначале Земля
имела первичную атмосферу. Но масса
юной Земли оказалась недостаточной
для удержания газов, и они
улетучивались. Земля увеличила
свою массу за счет космической пыли
и метеоритов: на Землю ежегодно
выпадает 107 кг пыли. К тому же Земля,
проходя через пылевое облако,
могла получать с космической пылью
10" т органического материала. Вторичная
атмосфера возникла тоже за счет дегазации
лав и состояла из СО, СОз, Нз, НзО, N, МНз.
Кислород появился в атмосфере благодаря
фотолизу - разложению паров воды в верхних
слоях атмосферы солнечными лучами. Позже
обогащение атмосферы кислородом шло
за счет фотосинтеза. Два с половиной миллиарда
лет назад исчезли золотоураносные конгломераты,
которые формируются только в отсутствии
кислорода. В тот же период появляются
красноцветы, образующиеся только при
наличии кислорода.
Второй этап химической
эволюции на Земле.
На этом этапе происходило
образование низкомолекулярных
органических соединений (аминокислот,
спиртов, углеводов, органических кислот).
Жизнь на Земле основана на углеродистых
соединениях. Почему именно углерод
стал основой жизни? Во-первых, потому,
что углерод образует соединения
в виде крупных молекулярных цепочек.
Во-вторых, углеродистые соединения взаимодействуют
медленно. В-третьих, углерод образует
сложные соединения с особой структурой,
существенной для протекания важнейших
жизненных процессов.
Химическая эволюция началась
задолго до возникновения Земли
- она началась в Космосе. В межзвездном
пространстве обнаружено более 50 органических
соединений. В Космосе обычен формальдегид,
окись углерода, вода, аммиак, цианистый
водород. Эти вещества, как показали эксперименты,
могут быть предшественниками аминокислот
и других органических соединений. Во
внеземном пространстве обнаружены углеводороды,
альдегиды, эфиры, аминокислоты, нуклеотиды,
ароматические соединения. Обнаружено
вещество, имеющее в своем составе 18 атомов
углерода. Синтез примитивных углеводородов,
начавшийся в Космосе, продолжался во
время формирования Солнечной системы
и Земли.
Предположения о процессах
второго этапа химической эволюции
имеют экспериментальное подтверждение.
В 1850 г. немецкий химик А. Штеккер осуществил
химический синтез аминокислот из аммиака,
альдегидов, синильной кислоты. В 1861 г.
А. М. Бутлеров, нагревая формальдегид
в крепком щелочном растворе, получил
смесь Сахаров. Д. И. Менделеев получал
углеводы, подвергая карбиды действию
водяного пара. Студент Чикагского университета
С. Л. Миллер в 1953 г. для дипломной работы,
выполненной под руководством С. Фокса,
собрал специальный аппарат для проверки
возможности абиогенетического синтеза
органических соединений. В этом герметическом
приборе в течение недели по замкнутой
схеме циркулировала смесь газов, которые,
по общему мнению, наиболее вероятно содержались
в ранней атмосфере Земли: СН4, Н, NH. Кипящая
вода - источник водяного пара - и холодильник
поддерживали циркуляцию газовой смеси.
В приборе непрерывно пропускали искры
при напряжении 60 тыс. вольт. После этого
воду подвергли хроматографическому и
химическому анализу. Было обнаружено
6 аминокислот (глицин, аланин, аспаргиновая
и глутаминовая кислоты и др.), мочевину,
молочную, янтарную, уксусную кислоты.
Всего было обнаружено 11 органических
кислот.
В том, что абиогенетический
синтез органики возможен, убеждает такой
факт: одно извержение вулкана в
настоящее время сопровождается
выбросом до 15 т органического вещества.
К тому же Земля, проходя через
пылевое облако, могла получать с космической
пылью 108 т органического материала. Все
это, предположительно, могло создать
тот "бульон", о котором писали А.
Опарин и Дж. Холдейн.
Начальные этапы биологической эволюции.
Образование первичных клеточных организмов положило начало биологической эволюции. Считается, что отбор коацерватов и пограничный этап химической и биологической эволюции продолжались около 750 млн лет. В конце этого периода появились первые примитивные безъядерные клетки - прокариоты. Первые живые организмы - гетеротрофы - использовали в качестве источника энергии (пищи) органические соединения, растворенные в водах первичного океана. Поскольку в атмосфере Земли не было свободного кислорода, гетеротрофы имели анаэробный (бескислородный) тип обмена веществ, эффективность которого невысока. Увеличение количества гетеротрофов привело к истощению вод первичного океана, где оставалось все меньше готовых органических веществ, которые можно было использовать для питания.В более выгодном положении оказались организмы, которые развили способность использовать энергию солнечного излучения для синтеза органических веществ из неорганических - фотосинтеза. Таким образом, появился принципиально новый источник питания. Например, современные фотосинтезирующие пурпурные бактерии благодаря солнечному излучению окисляют сероводород до сульфатов. Высвобождающийся в результате реакции окисления водород идет на восстановление диоксида углерода до углеводов с образованием воды. Использование органических соединений в качестве источника (донора) водорода привело к появлению автотрофных организмов (способных синтезировать из неорганических веществ все необходимые для жизни органические вещества).
Следующий шаг эволюции связан с развитием у фотосинтезирующих организмов способности использовать воду в качестве источника водорода для синтеза органических молекул. Усвоение углекислого газа такими организмами сопровождалось выделением кислорода и включением углерода в органические соединения. Так в атмосфере Земли начал накапливаться кислород. Первыми фотосинтезирующими организмами, выделяющими в атмосферу кислород, были цианобактерии (цианеи).
Переход от первичной атмосферы к среде, содержащей кислород, представляет собой важнейшее событие как в эволюции живых существ, так и в преобразовании минералов. Во - пеpвых, кислород, выделяющийся в атмосферу, в верхних ее слоях под действием мощного ультрафиолетового излучения Солнца превращается в активный озон (О3), который способен поглощать большую часть жестких коротковолновых ультрафиолетовых лучей, разрушительно действующих на сложные органические соединения. Во-вторых, в присутствии свободного кислорода возможен кислородный тип обмена веществ, энергетически более выгодный. Образование свободного кислорода вызвало к жизни многочисленные новые формы аэробных живых организмов и более широкое использование ими ресурсов окружающей среды.
В результате взаимополезного симбиоза различных прокариотических (не обладающих оформленным клеточным ядром) клеток возникли ядерные, или эукариотические, организмы (эукариоты). Основой симбиоза была, вероятно, гетеротрофная амебоподобная клетка. Питанием для нее служили более мелкие клетки и, в частности, дышащие кислородом аэробные бактерии, способные функционировать и внутри клетки-хозяина, производя энергию. Те крупные амебовидные клетки, в теле которых аэробные бактерии оставались невредимыми, оказались в более выгодном положении, чем клетки, получавшие энергию анаэробным путем - брожением. В дальнейшем бактерии-симбионты превратились в митохондрии (органеллы клеток, где протекают реакции, обеспечивающие клетки энергией). Когда к поверхности клетки-хозяина прикрепилась вторая группа симбионтов - жгутикоподобных бактерий, сходных с современными спирохетами, подвижность и способность к нахождению пищи такого организма резко возросли. Так возникли примитивные животные клетки - предшественники нынешних жгутиковых простейших.
Образовавшиеся подвижные эукариоты путем симбиоза с фотосинтезирующими (возможно, цианобактериями) организмами дали водоросль, или растение, причем строение пигментного комплекса у фотосинтезирующих анаэробных бактерий сходно с пигментами зеленых растений. Такое сходство указывает на возможность эволюционного преобразования фотосинтезирующего аппарата анаэробных бактерий в аналогичный аппарат зеленых растений.
Изложенную гипотезу о возникновении эукариотических клеток через ряд последовательных симбиозов приняли многие современные ученые, поскольку она хорошо обоснованна. Во-первых, одноклеточные водоросли и сейчас легко вступают в союз с животными - эукариотами; например, в теле инфузории туфельки обитает водоросль хлорелла. Во-вторых, некоторые органоиды клетки - митохондрии и пластиды - по строению ДНК очень похожи на прокариотические клетки-бактерии и цианобактерии.
Возможности эукариот по использованию среды существенно выше, чем у прокариот, поскольку они имеют диплоидный (двойной) набор генов. У прокариот любая мутация сразу проявляется в виде признака. Если мутация полезна, организм продолжает существовать, если вредна - он погибает, т.е. прокариоты непрерывно приспосабливаются к изменениям окружающей среды, но лишены возможности формировать крупные структурные изменения. Появление двойного набора генов у эукариот сделало возможным накопление непроявляющихся фенотипических мутаций и, следовательно, формирование резерва наследственной изменчивости - основы эволюционных преобразований.
Возможности одноклеточных в освоении среды обитания были ограничены, так как дыхание и питание простейших осуществляются через поверхность тела. При увеличении размеров клетки одноклеточного организма его поверхность возрастает по квадратичному закону, а объем - по кубическому, поэтому биологическая мембрана, окружающая клетку, не могла обеспечивать кислородом слишком большой организм. Иной эволюционный путь осуществился позже, около 2,6 млрд лет назад, когда появились многоклеточные организмы, эволюционные возможности которых значительно шире.
Первая гипотеза о происхождении многоклеточных организмов принадлежит Э. Геккелю (вторая половина XIX в.). При ее построении он исходил из исследований эмбрионального развития ланцетника (род животных класса бесчерепных), проведенных А.О. Ковалевским и другими зоологами. Геккель полагал, что начальная стадия развития зародыша (стадия зиготы) соответствует одноклеточным предкам, а стадия развития зародыша многоклеточных животных в процессе бластуляции (заключительной фазе периода дробления яйца) - шарообразной колонии жгутиковых. В дальнейшем, согласно этой гипотезе, произошло впячивание (инвагинация) одной из сторон шарообразной колонии и образовался гипотетический двухслойный организм, названный Геккелем гастреей. Теория Геккеля сыграла важную роль в истории науки, способствуя утверждению монофилетических (т.е. из одного корня) представлений о происхождении многоклеточных.
Основу современных представлений о возникновении многоклеточных организмов составляет гипотеза фагоцителлы И.И. Мечникова. По его представлениям, многоклеточные произошли от колониальных простейших - жгутиковых. Пример такой организации - ныне существующие колониальные жгутиковые типа вольвокс. Среди клеток колонии выделяются движущиеся, снабженные жгутиками, фагоцитирующие добычу и уносящие ее внутрь колонии, и половые, функцией которых является размножение. Так колония превратилась в примитивный, но целостный многоклеточный организм. О справедливости гипотезы фагоцителлы говорит строение примитивного многоклеточного организма - трихоплакса, который по строению соответствует гипотетической фагоцителле и поэтому должен быть выделен в особый тип животных - фагоцителлоподобных, заполняющих брешь между многоклеточными и одноклеточными организмами.
Таким образом, в настоящее время большинство исследователей в области естествознания признает, что возникновение жизни на Земле связано с длительным процессом химической эволюции. Формирование структуры, отграничивающей организм от окружающей среды, - мембраны с присущими ей свойствами способствовало появлению живых организмов и ознаменовало начало биологической эволюции. Как простейшие живые организмы, возникшие около 3 млрд лет назад, так и устроенные более сложно в основе своей структурной организации имеют клетку.
Основные направления
биологической эволюции.
В протерозойской эре в морях
обитало множество водорослей. Начальные
зве
и т.д.................
Химическая эволюция
Также этими терминами обозначается теория возникновения и развития тех молекул, которые имеют принципиальное значение для возникновения и развития живого вещества.
Всё, что известно о химизме вещества, позволяет ограничить проблему химической эволюции рамками так называемого «водно-углеродного шовинизма», постулирующего, что жизнь в нашей Вселенной представлена в единственно возможном варианте: в качестве «способа существования белковых тел», осуществимого благодаря уникальному сочетанию полимеризационных свойств углерода и деполяризующих свойств жидко-фазной водной среды, как совместно необходимых и/или достаточных(?) условий для возникновения и развития всех известных нам форм жизни. При этом подразумевается, что, по крайней мере, в пределах одной сформировавшейся биосферы может существовать только один, общий для всех живых существ данной биоты код наследственности, но пока остается открытым вопрос, существуют ли иные биосферы вне Земли и возможны ли иные варианты генетического аппарата.
Также неизвестно, когда и где началась химическая эволюция. Возможны любые сроки по окончании второго цикла звёздообразования, наступившего после конденсации продуктов взрывов первичных сверхновых звезд, поставляющих в межзвездное пространство тяжелые элементы (с атомной массой более 26). Второе поколение звёзд, уже с планетными системами, обогащенными тяжёлыми элементами, которые необходимы для реализации химической эволюции появилось через 0,5-1,2 млрд лет после Большого взрыва. При выполнении некоторых вполне вероятных условий, для запуска химической эволюции может быть пригодна практически любая среда: глубины океанов, недра планет, их поверхности, протопланетные образования и даже облака межзвёздного газа, что подтверждается повсеместным обнаружением в космосе методами астрофизики многих видов органических веществ - альдегидов, спиртов, сахаров и даже аминокислоты глицина, которые вместе могут служить исходным материалом для химической эволюции, имеющей своим конечным результатом возникновение жизни.
Методология исследования химической эволюции (теория)
Исследование химической эволюции осложняется тем, что в настоящее время знания о геохимических условиях древней Земли не являются достаточно полными.
Поэтому, кроме геологических, привлекаются также астрономические данные. Так, условия на Венере и Марсе рассматривают как близкие к тем, что были на Земле на различных этапах её эволюции.
Основные данные о химической эволюции получены в результате модельных экспериментов, в ходе которых удалось получить сложные органические молекулы при имитации различных химических составов атмосферы, гидросферы и литосферы и климатических условий.
На основе имеющихся данных был выдвинут ряд гипотез о конкретных механизмах и непосредственных движущих силах химической эволюции.
Абиогенез
Абиогене́з - образование органических соединений, распространённых в живой природе, вне организма без участия ферментов.
В широком смысле абиогенез - возникновение живого из неживого, то есть исходная гипотеза современной теории происхождения жизни
Существует также теория гиперциклов; согласно которой первые проявления жизни были соответственно в виде гиперциклов - комплекса сложных каталитических реакций, продукты выхода которых являются катализаторами для последующих реакций.
В 2008 году американские биологи сделали важный шаг к пониманию начальных этапов зарождения жизни. Им удалось создать «протоклетку» с оболочкой из простых липидов и жирных кислот, способную втягивать из окружающей среды активированные нуклеотиды - «кирпичики», необходимые для синтеза ДНК . В 2011 году японские ученые сообщили, что им удалось создать синтетическую клетку с оболочкой и элементами ДНК внутри, способную к размножению при нагревании "первичного бульона" до 94 градусов по Цельсию.
Эволюция
Биологическая эволюция - естественный процесс развития живой природы, сопровождающийся изменением генетического состава популяций, формированием адаптаций, видообразованием и вымиранием видов, преобразованием экосистем и биосферы в целом.
Существует несколько эволюционных теорий, объясняющих механизмы, лежащие в основе эволюционных процессов. В данный момент общепринятой является синтетическая теория эволюции (СТЭ), являющаяся развитием теории Дарвина. СТЭ позволяет объяснить связь субстрата эволюции (гены) и механизма эволюции (естественный отбор). В рамках СТЭ эволюция представляет собой процесс изменения наследственных черт в популяциях организмов в течение времени, превышающего продолжительность жизни одного поколения.
Чарльз Дарвин первым сформулировал теорию эволюции путем естественного отбора. Эволюция путем естественного отбора - это процесс, который следует из трех фактов о популяциях: 1) рождается больше потомства, чем может выжить; 2) у разных организмов разные черты, что приводит к различиям в выживаемости и вероятности оставить потомство; 3) эти черты - наследуемые. Таким образом, в следующем поколении увеличится количество таких особей, черты которых, способствуют выживанию и размножению в этой среде. Естественный отбор - единственная известная причина адаптаций, но не единственная причина эволюции. К числу неадаптивных причин относятся генетический дрейф, поток генов и мутации.
Несмотря на неоднозначное восприятие в обществе, факт эволюции является одним из наиболее доказанных в биологии. Открытия в эволюционной биологии оказали огромное влияние не только на традиционные области биологии, но и на другие академические дисциплины, такие как антропология и психология.
Введение
Эволюция происходит в течении периода времени превышающего срок жизни одного поколения и заключается в изменении наследуемых черт организма. Первым этапом этого процесса является изменение частот аллелей генов в популяции. В идеальной популяции, в которой отсутствует влияние внешней среды, дрейф и поток генов, согласно закону Харди-Вайнберга частота аллелей будет неизменна от поколения к поколению. Мутации увеличивают изменчивость в популяции за счет появления новых аллельных вариантов генов - мутационной изменчивости. Кроме мутационной, также есть комбинативная изменчивость, обусловленная рекомбинацией, но она не приводит не к изменениям частот аллелей, а к их новым сочетаниям. Ещё одним фактором приводящим к изменению частот аллелей является поток генов.
Два других фактора эволюции - естественный отбор и дрейф генов «сортируют» созданную мутациями и потоком генов изменчивость приводя к установлению новой частоты аллелей в популяции. Дрейф генов - вероятностный процесс изменения частот генов и наиболее ярко проявляется в популяциях относительно небольшого размера. Дрейф может приводить к полному исчезновению определенных аллелей из популяции. Естественный отбор является главным творческим фактором эволюции. Под его действием особи с определенным фенотипом (и определенным набором наследственных черт) будут более успешны чем другие, то есть будут иметь более высокую вероятность выжить и оставить потомство. Таким образом, в популяции будет увеличиваться доля таких организмов, у которых есть наследственные черты, обладающие селективным преимуществом. Взаимное влияние дрейфа и естественного отбора однозначно оценить сложно, но в целом, вероятно, оно зависит от размера популяции и интенсивности отбора. Кроме выше названных факторов важное значение может иметь и горизонтальный перенос генов, который может приводить к появлению совершенно новых, для данного организма генов.
Естественный отбор повышает приспособленность организмов, приводя к формированию адаптаций. Протекающие в течении длительного времени эволюционные процессы могут привести как к образованию новых видов и их дальнейшей дивергенции, так и к вымиранию целых видов.
Наследственность
Наследственность - это свойство организмов повторять в ряду поколений сходные типы обмена веществ и индивидуального развития в целом. Эволюция организмов происходит посредством изменения наследственных признаков организма. Примером наследственного признака у человека может служить коричневый цвет глаз, унаследованный от одного из родителей. Наследственные признаки контролируются генами. Совокупность всех генов организма образует его генотип.
Наследуемость также может встречаться и в более крупных масштабах. Например, экологическое наследование посредством строительства ниши. Таким образом, потомки наследуют не только гены, но и экологические особенности местообитания, созданного активностью из предков. Другими примерами наследования, не находящегося под контролем генов, может служить наследование культурных черт и симбиогенез.
Изменчивость
Фенотип организма обусловлен его генотипом и влиянием окружающей среды. Существенная часть вариаций фенотипов в популяциях вызвана в различиях их генотипов. В СТЭ эволюция определяется как изменение с течением времени генетической структуры популяций. Частота одной из аллелей изменяется, становясь более-менее распространенной среди других форм этого гена. Действующие силы эволюции ведут к изменениям в частоте аллели в одну или в другую стороны. Изменение исчезает, когда новая аллель достигает точки фиксации - целиком заменяет предковую аллель или исчезает из популяции.
Изменчивость складывается из мутаций, потока генов и рекомбинации генетического материала. Изменчивость также увеличивается за счет обменов генами между разными видами, таких как горизонтальный перенос генов у бактерий, гибридизация у растений. Несмотря на постоянные увеличение изменчивости за счет этих процессов, большая часть генома идентична у всех представителей данного вида. Однако даже сравнительно небольшие изменения в генотипе могут вызвать огромные различия в фенотипе, например, геномы шимпанзе и людей различаются всего на 5 %.
Мутации
Случайные мутации постоянно происходят в геномах всех организмов. Эти мутации создают генетическую изменчивость. Мутации - изменения в последовательности ДНК. Они вызваны радиацией, вирусами, транспозонами, мутагенными веществами, а также ошибками, происходящими во время репликации ДНК или мейоза. Мутации могут не иметь никакого эффекта, могут изменять продукт гена или препятствовать его функционированию. Исследования, проведенные на дрозофиле, показали, что, если мутация изменяет белок, производимый геном, то примерно в 70 % случаев это будет иметь вредные воздействия, а в остальных случаях нейтральные или слабоположительные. Для уменьшения негативного эффекта мутаций в клетках существует механизмы репарации ДНК. Оптимальный уровень мутаций - это баланс между высоким уровнем вредных мутаций и затратами на поддержание системы репарации. У РНК-вирусов уровень мутабильности высокий, что, видимо, является преимуществом, помогая избегать защитных ответов иммунной системы.
Мутации могут вовлекать большие участки хромосом. Например, при дупликации, которая вызывает появление дополнительных копий гена в геноме. Эти копии становятся основным материалом для возникновения новых генов. Это важный процесс, поскольку новые гены развиваются в пределах семейства генов от общего предка.
Рекомбинация
У бесполых организмов гены во время размножения не могут смешиваться с генами других особей. В отличие от них, у организмов с половым размножением потомство получает случайные смеси хромосом своих родителей. Это происходит за счет процесса гомологичной рекомбинации в ходе которого происходит обмен участков двух гомологичных хромосом. При рекомбинации не происходит изменения частоты аллелей, но происходит образование их новых комбинаций. Таким образом, половое размножение обычно увеличивает наследственную изменчивость и может ускорять темп эволюции организма. Однако, бесполое размножение зачастую выгодно и может развиваться у животных с половым размножением. Это может позволить двум наборам аллелей в геноме дивергировать с приобретением новых функций.
Рекомбинация позволяет наследоваться независимо даже аллелям, которые находятся близко друг от друга в ДНК. Однако, уровень рекомбинации низок - примерно две рекомбинации на хромосому за поколение.
Поток генов
Потоком генов называют перенос аллелей генов между популяциями. Поток генов может осуществляться за счет миграций особей между популяциями в случае подвижных организмов, либо, например, с помощью переноса пыльцы или семян в случае растений. Скорость потока генов сильно зависит от подвижности организмов.
Степень влияние потока генов на изменчивость в популяциях до конца не ясна. Существуют две точки зрения, одна из них, что поток генов может иметь значительное влияние на крупных популяционных системах, гомогенизируя их и, соответственно, действую против процессов видообразования; вторая, что скорости потока генов достаточно, только для воздействия на локальные популяции.
Механизмы эволюции
Естественный отбор
Эволюция посредством естественного отбора - это процесс, при котором закрепляются мутации, увеличивающие приспособленность организмов. Естественный отбор часто называют «самоочевидным» механизмом, поскольку следует из таких фактов как:
- Наследственные изменения существуют в популяциях организмов;
- Организмы производят потомков больше, чем может выжить;
- Эти потомки различаются тем, что имеют различную выживаемость и способность размножаться.
Такие условия создают конкуренцию между организмами в выживании и размножении. Таким образом, организмы с наследственными чертами, которые дают им конкурентное преимущество, имеют большую вероятность передать их своим потомкам, чем организмы с наследственными чертами, не имеющими подобного преимущества.
Центральное понятие концепции естественного отбора - приспособленность организмов. Приспособленность определяется как способность организма к выживанию и размножению, которая определяет размер его генетического вклада в следующее поколение. Однако, главным в определении приспособленности является не общее число потомков, а число потомков с данным генотипом (относительная приспособленность). Естественный отбор для черт, которые могут изменяться в некотором диапазоне значений (например, размер организма), можно разделить на три типа:
Описание работы
Химическая эволюции или пребиотическая эволюция - первый этап эволюции жизни, в ходе которого органические, пребиотические вещества возникли из неорганических молекул под влиянием внешних энергетических и селекционных факторов и в силу развертывания процессов самоорганизации, свойственных всем относительно сложным системам, которыми бесспорно являются все углерод-содержащие молекулы.
По мнению большинства ученых (в первую очередь астрономов и геологов), Земля сформировалась как небесное тело около 5 млрд лет т.н. путем конденсации частиц вращавшегося вокруг Солнца газопылевого облака.
Под влиянием сил сжатия частицы, из которых формируется Земля, выделяют огромное количество тепла. В недрах Земли начинаются термоядерные реакции. В результате Земля сильно разогревается. Таким образом, 5 млрд лет т.н. Земля представляла собой несущийся в космическом пространстве раскаленный шар, температура поверхности которою достигала 4000-8000°С (рис. 2.4.1.1).
Постепенно, за счет излучения тепловой энергии в космическое пространство, Земля начинает остывать. Около 4 млрд лет т.н. Земля остывает настолько, что на ее поверхности формируется твердая кора; одновременно из ее недр вырываются легкие, газообразные вещества, поднимающиеся вверх и формирующие первичную атмосферу. По составу первичная атмосфера существенно отличалась от современной. Свободный кислород в атмосфере древней Земли, по-видимому, отсутствовал, а в ее состав входили вещества в восстановленном состоянии, такие, как водород (Н 2), метан (СН 4), аммиак (NH 3), пары воды (Н 2 О), а возможно, также азот (N 2), окись и двуокись углерода (СО и С0 2).
Восстановительный характер первичной атмосферы Земли чрезвычайно важен для зарождения жизни, поскольку вещества в восстановленном состоянии обладают высокой реакционной способностью и в определенных условиях способны взаимодействовать друг с другом, образуя органические молекулы. Отсутствие в атмосфере первичной Земли свободного кислорода (практически весь кислород Земли был связан в виде окислов) также является важной предпосылкой возникновения жизни, поскольку кислород легко окисляет и тем самым разрушает органические соединения. Поэтому при наличии в атмосфере свободного кислорода накопление на древней Земле значительного количества органических веществ было бы невозможно.
Около 5 млрд лет т.п. - возникновение Земли как небесного тела; температура поверхности - 4000-8000°С
Около 4 млрд лет т.н. - формирование земной коры и первичной атмосферы
При температуре 1000°С - в первичной атмосфере начинается синтез простых органических молекул
Энергию для синтеза дают:
Температура первичной атмосферы ниже 100°С - формирование первичного океана -
Синтез сложных органических молекул - биополимеров из простых органических молекул:
простые органические молекулы - мономеры
сложные органические молекулы - биополимеры
Рис. 2.1. Основные этапы химической эволюции
Когда температура первичной атмосферы достигает 1000°С, в ней начинается синтез простых органических молекул, таких, как аминокислоты, нуклеотиды, жирные кислоты, простые сахара, многоатомные спирты, органические кислоты и др. Энергию для синтеза поставляют грозовые разряды, вулканическая деятельность, жесткое космическое излучение и, наконец, ультрафиолетовое излучение Солнца, от которого Земля еще не защищена озоновым экраном, причем именно ультрафиолетовое излучение ученые считают основным источником энергии для абиогенного (т.е. проходящею без участия живых организмов) синтеза органических веществ.
Признанию и широкому распространению теории А.И. Опарина во многом способствовало то, что процессы абиогенного синтеза органических молекул легко воспроизводятся в модельных экспериментах.
Возможность синтеза органических веществ из неорганических была известна с начала 19 в. Уже в 1828 г. выдающийся немецкий химик Ф. Вёлер синтезировал органическое вещество - мочевину из неорганическою - циановокислого аммония. Однако возможность абиогенного синтеза органических веществ в условиях, близких к условиям древней Земли, была впервые показана в опыте С. Миллера.
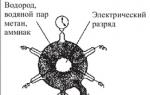
В 1953 г. молодой американский исследователь, студент- дипломник Чикагского университета Стенли Миллер воспроизвел в стеклянной колбе с впаянными в нес электродами первичную атмосферу Земли, которая, по мнению ученых того времени, состояла из водорода метана СН 4 , аммиака NH, и паров воды Н 2 0 (рис. 2.4.1.2). Через эту газовую смесь С. Миллер в течение недели пропускал электрические разряды, имитирующие грозовые. По окончании эксперимента в колбе были обнаружены α-аминокислоты (глицин, аланин, аспарагин, глутамин), органические кислоты (янтарная, молочная, уксусная, гликоколовая), у-оксимасляная кислота и мочевина. При повторении опыта С. Миллеру удалось получить отдельные нуклеотиды и короткие полинуклеотидные цепочки из пяти-шести звеньев.

Рис. 2.2. Установка С. Миллера
В дальнейших опытах по абиогенному синтезу, проводимых различными исследователями, использовались не только электрические разряды, но и другие виды энергии, характерные для древней Земли, - космическое, ультрафиолетовое и радиоактивное излучения, высокие температуры, присущие вулканической деятельности, а также разнообразные варианты газовых смеси, имитирующих первичную атмосферу. В результате был получен практически весь спектр органических молекул, характерных для живого: аминокислоты, нуклеотиды, жироподобные вещества, простые сахара, органические кислоты.
Более того, абиогенный синтез органических молекул может происходить на Земле и в настоящее время (например, в процессе вулканической деятельности). При этом в вулканических выбросах можно обнаружить не только синильную кислоту HCN, являющуюся предшественником аминокислот и нуклеотидов, но и отдельные аминокислоты, нуклеотиды и даже такие сложные по строению органические вещества, как порфирины. Абиогенный синтез органических веществ возможен не только на Земле, но и в космическом пространстве. Простейшие аминокислоты обнаружены в составе метеоритов и комет.
Когда температура первичной атмосферы опустилась ниже 100°С, на Землю обрушились горячие дожди и появился первичный океан. С потоками дождя в первичный океан поступали абиогенно синтезированные органические вещества, что превратило его, но образному выражению английского биохимика Джона Холдейна, в разбавленный «первичный бульон». По-видимому, именно в первичном океане начинаются процессы образования из простых органических молекул - мономеров сложных органических молекул - биополимеров (см. рис. 2.4.1.1).
Однако процессы полимеризации отдельных нуклеогидов, аминокислот и Сахаров - это реакции конденсации, они протекают с отщеплением воды, следовательно, водная среда способствует не полимеризации, а, напротив, гидролизу биополимеров (т.е. разрушению их с присоединением воды).
Образование биополимеров (в частности, белков из аминокислот) могло происходить в атмосфере при температуре около 180°С, откуда они смывались в первичный океан с атмосферными осадками. Кроме того, возможно, на древней Земле аминокислоты концентрировались в пересыхающих водоемах и полимеризовались в сухом виде под действием ультрафиолетового света и тепла лавовых потоков.
Несмотря на то что вода способствует гидролизу биополимеров, в живой клетке синтез биополимеров осуществляется именно в водной среде. Этот процесс катализируют особые белки-катализаторы - ферменты, а необходимая для синтеза энергия выделяется при распаде аденозинтрифосфорной кислоты - АТФ. Возможно, синтез биополимеров в водной среде первичного океана катализировался поверхностью некоторых минералов. Экспериментально показано, что раствор аминокислоты аланина может полимеризоваться в водной среде в присутствии особого вида глинозема. При этом образуется пептид полиаланин. Реакция полимеризации аланина сопровождается распадом АТФ.
Полимеризация нуклеотидов проходит легче, чем полимеризация аминокислот. Показано, что в растворах с высокой концентрацией солей отдельные нуклеотиды самопроизвольно полимеризуются, превращаясь в нуклеиновые кислоты.
Жизнь всех современных живых существ - это процесс непрерывного взаимодействия важнейших биополимеров живой клетки - белков и нуклеиновых кислот.
Белки - это «молекулы-рабочие», «молекулы-инженеры» живой клетки. Характеризуя их роль в обмене веществ, биохимики часто используют такие образные выражения, как «белок работает», «фермент ведет реакцию».Важнейшая функция белков- каталитическая . Как известно, катализаторы - это вещества, которые ускоряют химические реакции, но сами в конечные продукты реакции не входят.Бачки-катализаторы называются ферментами. Ферменты в согни и тысячи раз ускоряют реакции обмена веществ. Обмен веществ, а значит, и жизнь без них невозможны.
Нуклеиновые кислоты - это «молекулы-компьютеры», молекулы - хранители наследственной информации. Нуклеиновые кислоты хранят информацию не обо всех веществах живой клетки, а только о белках. Достаточно воспроизвести в дочерней клетке белки, свойственные материнской клетке, чтобы они точно воссоздали все химические и структурные особенности материнской клетки, а также свойственный ей характер и темпы обмена веществ. Сами нуклеиновые кислоты также воспроизводятся благодаря каталитической активности белков.
Таким образом, тайна зарождения жизни - это тайна возникновения механизма взаимодействия белков и нуклеиновых кислот. Какими же сведениями об этом процессе располагает современная наука? Какие молекулы явились первичной основой жизни - белки или нуклеиновые кислоты?
Ученые полагают, что несмотря на ключевую роль белков в обмене веществ современных живых организмов, первыми «живыми» молекулами были не белки, а нуклеиновые кислоты, а именно рибонуклеиновые кислоты (РНК).
В 1982 г. американский биохимик Томас Чек открыл автокаталитические свойства РНК. Он экспериментально показал, что в среде, содержащей в высокой концентрации минеральные соли, рибонуклеотиды спонтанно (самопроизвольно) полимеризуются, образуя полинуклеотиды - молекулы РНК. На исходных поли- нуклеотидных цепях РНК, как на матрице, путем спаривания комплементарных азотистых оснований образуются РНК-ко- пии. Реакция матричного копирования РНК катализируется исходной молекулой РНК и не требует участия ферментов либо других белков.
Дальнейшие события достаточно хорошо объясняются процессом, который можно было бы назвать «естественным отбором» на уровне молекул. При самокопировании (самосборке) молекул РНК неизбежно возникают неточности, ошибки. Содержащие ошибки копии РНК снова копируются. При повторном копировании вновь могут возникнуть ошибки. В результате популяция молекул РНК на определенном участке первичного океана будет неоднородна.
Поскольку параллельно с процессами синтеза идут и процессы распада РНК, в реакционной среде будут накапливаться молекулы, обладающие либо большей стабильностью, либо лучшими автокаталитическими свойствами (т.е. молекулы, которые быстрее себя копируют, быстрее «размножаются»).
На некоторых молекулах РНК, как на матрице, может происходить самосборка небольших белковых фрагментов - пептидов. Вокруг молекулы РНК образуется белковый «чехол».
Наряду с автокаталитическими функциями Томас Чек обнаружил у молекул РНК и явление самосплайсинга. В результате самосплайсинга участки РНК, не защищенные пептидами, самопроизвольно удаляются из РНК (они как бы «вырезаются» и «выбрасываются»), а оставшиеся участки РНК, кодирующие белковые фрагменты, «срастаются», т.е. самопроизвольно объединяются в единую молекулу. Эта новая молекула РНК уже будет кодировать большой сложный белок (рис. 2.4.1.3).
По-видимому, первоначально белковые чехлы выполняли в первую очередь, защитную функцию, предохраняя РНК от разрушения и повышая тем самым ее стабильность в растворе (такова функция белковых чехлов и у простейших современных вирусов).
Очевидно, что на определенном этапе биохимической эволюции преимущество получили молекулы РНК, кодирующие не только защитные белки, но и белки-катализаторы (ферменты), резко ускоряющие скорость копирования РНК. По-видимому, именно таким образом и возник процесс взаимодействия белков и нуклеиновых кислот, который мы в настоящее время называем жизнью.
В процессе дальнейшего развития, благодаря появлению белка с функциями фермента - обратной транскриптазы, на одно- цепочечных молекулах РНК стали синтезироваться состоящие из двух цепей молекулы дезоксирибонуклеиновой кислоты (ДНК). Отсутствие у дезоксирибозы ОН-группы в 2" положении делает молекулы ДНК более стабильными по отношению к гидролитическому расщеплению в слабощелочных растворах, а именно слабощелочной была реакция среды в первичных водоемах (эта реакция среды сохранилась и в цитоплазме современных клеток).
Где же происходило развитие сложного процесса взаимодействия белков и нуклеиновых кислот? По теории А.И. Опарина, местом зарождения жизни стали так называемые коацерватные капли.

Рис. 2.3.Гипотеза возникновения взаимодействия белков и нуклеиновых кислот:
а) в процессе самокопирования РНК накапливаются ошибки (1 - нуклеотиды, соответствующие исходной РНК; 2 - нуклеотиды, не соответствующие исходной РНК, - ошибки в копировании); б) на часть молекулы РНК за счет ее физико-химических свойств «налипают» аминокислоты (3 - молекула РНК; 4 - аминокислоты), которые, взаимодействуя друг с другом, превращаются в короткие белковые молекулы - пептиды.
В результате свойственного молекулам РНК самосплайсинга незащищенные пептидами участки молекулы РНК разрушаются, а оставшиеся «срастаются» в единую молекулу, кодирующую крупный белок.
В результате возникает молекула РНК, покрытая белковым чехлом (сходное строение имеют и наиболее примитивные современные вирусы, например вирус табачной мозаики)
Явление коацервации состоит в том, что в некоторых условиях (например, в присутствии электролитов) высокомолекулярные вещества отделяются от раствора, но не в форме осадка, а в виде более кон центрирован но го раствора - коацервата. При встряхивании коацерват распадается на отдельные мелкие капельки. В воде такие капли покрываются стабилизирующей их гидратной оболочкой (оболочкой из молекул воды) - рис. 2.4.1.4.
Коацерватные капли обладают некоторым подобием обмена веществ: иод воздействием чисто физико-химических сил они могут избирательно впитывать из раствора некоторые вещества и выделять в окружающую среду продукты их распада. За счет избирательного концентрирования веществ из окружающей среды они могут расти, а но достижении определенного размера начинают «размножаться», отпочковывая маленькие капельки, которые, в свою очередь, могут расти и «почковаться».
Возникшие в результате концентрирования белковых растворов коацерватные капли в процессе перемешивания под действием волн и ветра могут покрываться оболочкой из липи- дов: одинарной, напоминающей мицеллы мыла (при однократном отрыве капли от поверхности воды, покрытой липидным слоем), либо двойной, напоминающей клеточную мембрану (при повторном падении капли, покрытой однослойной липидной мембраной, на липидную пленку, покрывающую поверхность водоема - рис. 2.4.1.4).
Процессы возникновения коацерватных капель, их роста и «почкования», а также «одевания» их мембраной из двойного липидного слоя легко моделируются в лабораторных условиях.
Для коацерватных капель также существует процесс «естественного отбора», при котором в растворе сохраняются наиболее стабильные капли.
Несмотря на внешнее сходство коацерватных капель с живыми клетками, у коацерватных капель отсутствует главный признак живого - способность к точному самовоспроизведению, самокопированию. Очевидно, предшественниками живых клеток явились такие коацерватные капли, в состав которых вошли комплексы молекул-репликаторов (РНК или ДНК) и кодируемых ими белков. Возможно, комплексы РНК-белок длительное время существовали вне коацерватных капель в виде так называемого «свободноживущего гена», а возможно, их формирование проходило непосредственно внутри некоторых коацерватных капель.

Рис 2.4.Возможный путь перехода от коацерватных капель к примитивным клешам:
а) образование коацсрвата; 6) стабилизация коацерватных капель в водном растворе; в) - формирование вокруг капли двойного липидного слоя, похожего на клеточную мембрану: 1 - коацерватная капля; 2 - мономолекулярный слой липида на поверхности водоема; 3 - формирование вокруг капли одинарного липидного слоя; 4 - формирование вокруг капли двойного липидного слоя, похожего на клеточную мембрану; г) - коацерватная капля, окруженная двойным липидным слоем, с вошедшим в ее состав белково-нуклеотидным комплексом - прообраз первой живой клетки
Исключительно сложный, не до конца понятный современной науке процесс возникновения жизни на Земле прошел с исторической точки зрения чрезвычайно быстро. Уже 3,5 млрд лет т.н. химическая эволюция завершилась появлением первых живых клеток и началась биологическая эволюция.
Теория химической эволюции
(пребиотическая эволюция
, теория абиогенеза
) — первый этап эволюции жизни, в ходе которого органические, пребиотические вещества возникли из неорганических молекул под влиянием внешних энергетических и селекционных факторов и в силу развертывания процессов самоорганизации, свойственных всем относительно сложным системам, которыми бесспорно являются все углерод-содержащие молекулы. Также этими терминами обозначается теория возникновения и развития тех молекул, которые имеют принципиальное значение для возникновения и развития живого вещества.
Жизнь в нашей Вселенной представлена в единственно возможном варианте: в качестве «способа существования белковых тел», осуществимого благодаря уникальному сочетанию полимеризационных свойств углерода и деполяризующих свойств жидко-фазной водной среды, как совместно необходимых и достаточных условий для возникновения и развития всех известных нам форм жизни. При этом подразумевается, что, по крайней мере, в пределах одной сформировавшейся биосферы может существовать только один, общий для всех живых существ данной биоты код наследственности, но пока остается открытым вопрос, существуют ли иные биосферы вне Земли и возможны ли иные варианты генетического аппарата.
Исследования
Исследование химической эволюции осложняется тем, что в настоящее время знания о геохимических условиях древней Земли не являются достаточно полными. Поэтому, кроме геологических, привлекаются также астрономические данные. Так, условия на Венере и Марсе рассматривают как близкие к тем, что были на Земле на различных этапах её эволюции. Основные данные о химической эволюции получены в результате модельных экспериментов, в ходе которых удалось получить сложные органические молекулы при имитации различных химических составов атмосферы, гидросферы и литосферы и климатических условий. На основе имеющихся данных был выдвинут ряд гипотез о конкретных механизмах и непосредственных движущих силах химической эволюции.
Абиогенез
В широком смысле абиогенез — возникновение живого из неживого, то есть исходная гипотеза современной теории происхождения жизни. В 20-х годах XX века академик Александр Опарин предположил, что в растворах высокомолекулярных соединений могут самопроизвольно образовываться зоны повышенной концентрации, которые относительно отделены от внешней среды и могут поддерживать обмен с ней. Он назвал их коацерватные капли, или просто коацерваты .
В 1953 году Стэнли Миллером экспериментально осуществлён абиогенный синтез аминокислот и других органических веществ в условиях, воспроизводящих условия первобытной Земли. Существует также теория гиперциклов, согласно которой первые проявления жизни были соответственно в виде гиперциклов — комплекса сложных каталитических реакций, продукты выхода которых являются катализаторами для последующих реакций.
В 2008 году американские биологи сделали важный шаг к пониманию начальных этапов зарождения жизни. Им удалось создать «протоклетку» с оболочкой из простых липидов и жирных кислот, способную втягивать из окружающей среды активированные нуклеотиды — «кирпичики», необходимые для синтеза ДНК.
Аспекты
Гипотезы химической эволюции должны объяснять различные аспекты:
1.
Небиологическое начало биомолекул, то есть их развитие из неживущих и, соответственно, неорганических предшественников.
2.
Появление способных к саморепликации и самоизменению химических информационных систем, то есть возникновение клетки.
3.
Появление взаимной зависимости функции (ферментов) и информации (РНК, ДНК).
4.
Условия среды Земли в период от 4,5 до 3,5 млрд лет назад.
Унифицированная модель химической эволюции ещё не разработана, возможно потому, что основные принципы ещё не открыты.
Рассуждения
Биомолекулы
Пребиотический синтез сложных соединений молекул может делится на три последовательных этапа:
1.
Возникновение простых органических соединений (спиртов, кислот, гетероциклических соединений: пуринов, пиримидинов и пирролов) из неорганических материалов.
2.
Синтез более сложных органических соединений - "биомолекул" - представителей наиболее распространённых классов метаболитов, в том числе и мономеров - структурных единиц биополимеров (моносахаридов, аминокислот, жирных кислот, нуклеотидов) из простых органических соединений.
3.
Возникновение сложных биополимеров (полисахариды, белки, нуклеиновые кислоты) из основных структурных единиц - мономеров.
Развитие древней атмосферы
Развитие земной атмосферы является частью химической эволюции и к тому же важным элементом истории климата. Сегодня её разделяют на четыре важные ступени развития.
В начале происходило образование химических элементов в космосе и появление из них земли — приблизительно 4,56 миллиардов лет назад. Предположительно наша планета уже довольно рано имела атмосферу из водорода и гелия, которая однако постепенно утекла в космическое пространство. Астрономы исходят так же из того, что из-за относительно высоких температур и эффектов солнечного ветра на Земле и на других близлежащих к Солнцу планетах могло остаться небольшое количество легких химических элементов (включая углерод, азот и кислород). Все эти элементы, составляющие сегодня основную часть биосферы, были занесены ударами комет из внешних участков солнечной системы лишь через большой промежуток времени, когда протопланеты немного остыли. В течение первых нескольких миллионов лет после возникновения солнечной системы постоянно повторялись столкновения с небесными телами, а вызванные ими коллизии уничтожали образованные в это время живые системы. Поэтому появление жизни смогло начаться только после накопления воды за длительное время хотя бы в самых глубоких впадинах.
С медленным остыванием земли, вулканической деятельностью (выделение газов из недр земли) и глобальным распределением материалов упавших комет возникла вторая атмосфера земли. Скорее всего она состояла из водяного пара (H2O до 80 %), углекислого газа (CO2 до 20 %), сероводорода (до 7 %), аммиака и метана. Высокий процент водяного пара объясняется тем, что поверхность земли была на тот момент ещё слишком горяча для образования морей. Прежде всего из воды, метана и аммиака в условиях молодой земли могли образоваться небольшие органические молекулы (кислоты, спирты, аминокислоты), позднее - органические полимеры (полисахариды, жиры, полипептиды), которые были нестабильны в кислотной атмосфере.
После охлаждения атмосферы до температуры ниже кипения воды наступил очень длительный дождь, который и образовал океаны. Насыщенность других газов атмосферы относительно водяного пара повысилась. Высокое ультрафиолетовое облучение обусловило фотохимический распад воды, метана и аммиака, в результате чего накопились углекислый газ и азот. Лёгкие газы — водород и гелий — уносились в космос, углекислый газ растворялся в больших количествах в океане, окисляя воду. Значение pH упало до 4. Инертный и малорастворимый азот N2 накапливался со временем и образовал около 3,4 миллиардов лет назад основную составляющую атмосферы.
Выпадание в осадок прореагировавшего с ионами металлов растворенного углекислого газа (карбонаты) и дальнейшее развитие живых существ, которые ассимилировали углекислый газ, привело к уменьшению CO2-концентрации и поднятию значения pH в водоёмах.
Кислород O2 играл важнейшую роль в дальнейшем развитии атмосферы. Он образовался с появлением способных к фотосинтезу живых существ, предположительно цианобактерий (сине-зелёных водорослей) или им подобных прокариотов. Ассимиляция ими углекислого газа привела к дальнейшему понижению кислотности, но насыщенность атмосферы кислородом оставалась все-таки довольно низкой. Причина этого — незамедлительное использование растворенного в океане кислорода для окисления двухвалентных ионов железа и других окисляемых соединений. Около двух миллиардов лет назад этот процесс завершился, и кислород стал постепенно накапливаться в атмосфере.
Очень реактивный кислород легко окисляет восприимчивые органические биомолекулы и становится таким образом фактором отбора окружающей среды для ранних организмов. Только немногие анаэробные организмы смогли переместиться в свободные от кислорода жизненные пространства, другая часть выработала ферменты, которые делают кислород не опасным.
Миллиард лет назад содержание кислорода в атмосфере перешагнуло планку одного процента и спустя несколько миллионов лет был образован озоновый слой. Сегодняшнее содержание кислорода в 21 % было достигнуто лишь 350 миллионов лет назад и сохраняется с тех пор стабильным.
Значение воды для возникновения жизни
H2O — химическое соединение, присутствующие при обычных условиях во всех трёх агрегатных состояниях.
Для жизни, как мы её знаем (или определяем), нужна вода как универсальный растворитель. Вода имеет комплекс качеств, которые делают жизнь возможной. Нет данных, что жизнь может возникать и существовать независимо от воды и принято считать, что только наличие воды в жидкой фазе (в определённой области или на определённой планете), делает вероятным возникновение там жизни.
Иллюстрации


Рис. 1-2. Наземный и глубоководный вулканы - вероятные предусловия возникновения жизни на Земле
Единство происхождения жизни на Земле и причины неоднородности и разнообразия живых организмов
Выполнила:
студентка естественно –
геграфического факультета
гр. БИ – 11
Фролова Алла Александровна
Ульяновск, 2014 г
Глава I. Единство происхождения. 3
1. 1. Предбиологическая (химическая) эволюция. 3
1. 2. Основные этапы химической эволюции. 3
Глава II. Причины неоднородности и разнообразия. 7
Используемая литература. 10
Глава I. Единство происхождения.
Предбиологическая (химическая) эволюция.
По мнению большинства ученых (в первую очередь астрономов и геологов), Земля сформировалась как небесное тело около 5 млрд лет т.н. путем конденсации частиц вращавшегося вокруг Солнца газопылевого облака.
Восстановительный характер первичной атмосферы Земли чрезвычайно важен для зарождения жизни, поскольку вещества в восстановленном состоянии в определенных условиях способны взаимодействовать друг с другом, образуя органические молекулы. Отсутствие в атмосфере первичной Земли свободного кислорода (практически весь кислород Земли был связан в виде окислов) также является важной предпосылкой возникновения жизни, поскольку кислород легко окисляет и тем самым разрушает органические соединения. Поэтому при наличии в атмосфере свободного кислорода накопление на древней Земле значительного количества органических веществ было бы невозможно.
Основные этапы химической эволюции.
Когда температура первичной атмосферы достигает 1000°С, в ней начинается синтез простых органических молекул, таких, как аминокислоты, нуклеотиды, жирные кислоты, простые сахара, многоатомные спирты, органические кислоты и др. Энергию для синтеза поставляют грозовые разряды, вулканическая деятельность, жесткое космическое излучение и, наконец, ультрафиолетовое излучение Солнца, от которого Земля еще не защищена озоновым экраном.
Когда температура первичной атмосферы опустилась ниже 100°С, на Землю обрушились горячие дожди и появился первичный океан. С потоками дождя в первичный океан поступали абиогенно синтезированные органические вещества, что превратило его, в разбавленный «первичный бульон». По-видимому, именно в первичном океане начинаются процессы образования из простых органических молекул - мономеров сложных органических молекул - биополимеров.
Образование биополимеров (в частности, белков из аминокислот) могло происходить и в атмосфере при температуре около 180°С. Кроме того, возможно, на древней Земле аминокислоты концентрировались в пересыхающих водоемах и полимеризовались в сухом виде под действием ультрафиолетового света и тепла лавовых потоков.
Полимеризация нуклеотидов проходит легче, чем полимеризация аминокислот. Показано, что в растворах с высокой концентрацией солей отдельные нуклеотиды самопроизвольно полимеризуются, превращаясь в нуклеиновые кислоты.
Жизнь всех современных живых существ - это процесс непрерывного взаимодействия важнейших биополимеров живой клетки - белков и нуклеиновых кислот.
Таким образом, тайна зарождения жизни - это тайна возникновения механизма взаимодействия белков и нуклеиновых кислот.
Где же происходило развитие сложного процесса взаимодействия белков и нуклеиновых кислот? По теории А.И. Опарина, местом зарождения жизни стали так называемые коацерватные капли.
Гипотеза возникновения взаимодействия белков и нуклеиновых кислот:
Явление коацервации состоит в том, что в некоторых условиях (например, в присутствии электролитов) высокомолекулярные вещества отделяются от раствора, но не в форме осадка, а в виде более концентрированного раствора - коацервата. При встряхивании коацерват распадается на отдельные мелкие капельки. В воде такие капли покрываются стабилизирующей их гидратной оболочкой (оболочкой из молекул воды) - рис. 2.4.1.4.
Коацерватные капли обладают некоторым подобием обмена веществ: под воздействием чисто физико-химических сил они могут избирательно впитывать из раствора некоторые вещества и выделять в окружающую среду продукты их распада. За счет избирательного концентрирования веществ из окружающей среды они могут расти, а но достижении определенного размера начинают «размножаться», отпочковывая маленькие капельки, которые, в свою очередь, могут расти и «почковаться».
Возникшие в результате концентрирования белковых растворов коацерватные капли в процессе перемешивания под действием волн и ветра могут покрываться оболочкой из липидов: одинарной, напоминающей мицеллы мыла (при однократном отрыве капли от поверхности воды, покрытой липидным слоем), либо двойной, напоминающей клеточную мембрану (при повторном падении капли, покрытой однослойной липидной мембраной, на липидную пленку, покрывающую поверхность водоема).
Процессы возникновения коацерватных капель, их роста и «почкования», а также «одевания» их мембраной из двойного липидного слоя легко моделируются в лабораторных условиях.
Для коацерватных капель также существует процесс «естественного отбора», при котором в растворе сохраняются наиболее стабильные капли.
Несмотря на внешнее сходство коацерватных капель с живыми клетками, у коацерватных капель отсутствует главный признак живого - способность к точному самовоспроизведению, самокопированию. Очевидно, предшественниками живых клеток явились такие коацерватные капли, в состав которых вошли комплексы молекул-репликаторов (РНК или ДНК) и кодируемых ими белков. Возможно, комплексы РНК-белок длительное время существовали вне коацерватных капель в виде так называемого «свободноживущего гена», а возможно, их формирование проходило непосредственно внутри некоторых коацерватных капель.
Исключительно сложный, не до конца понятный современной науке процесс возникновения жизни на Земле прошел с исторической точки зрения чрезвычайно быстро. Уже 3,5 млрд лет т.н. химическая эволюция завершилась появлением первых живых клеток и началась биологическая эволюция. (URL:http://www.grandars.ru/shkola/geografiya/proishozhdenie-zhizni (дата обращения: 28.09.2014).